האוויר שאנו נושמים מכיל כ-78% חנקן, 21% חמצן, 0.3% פחמן דו-חמצני וגזים אחרים בכמויות קטנות. החמצן הוא גורם מפתח במטבוליזם שלנו,
ובלעדיו איננו יכולים להתקיים.
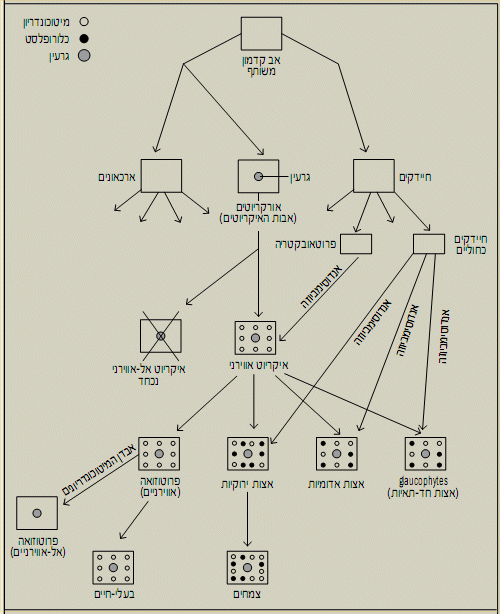
ביצורים האיקריוטיים (Eukarya - בעלי הגרעין), האברון התוך-תאי ה"מטפל" בחמצן ומאפשר את השימוש בו לצרכים אנרגטיים הוא המיטוכונדריון.
בממוצע יש כ-50 מיטוכונדריונים בכל תא, ובהם מתקיימים תהליכי המפתח של המטבוליזם החמצני בתא -מעגל קרבס ושרשרת העברת האלקטרונים שבה נצרך
החמצן ונפלטים מים (הנשימה החמצנית).

החיים בנוכחות חמצן נראים לנו כדבר המובן מאליו. אך לא תמיד זה היה כך. כשהופיעו לראשונה החיים על פני כדור-הארץ, לפני כ-3.8 מיליארדי
שנה, הרכב האטמוספרה היה שונה לגמרי - היא הכילה מֶתאן (CH4), חנקן, פחמן דו-חמצני, מימן וגזים נוספים, אך לא חמצן. היצורים
הראשונים שהופיעו על פני כדור-הארץ היו חיידקים שהיום אנו מגדירים אותם כאל-אווירניים (anaerobes; האפיון "אווירני" מתייחס לחלק החמצני
של האוויר). חיידקים אלה התפצלו לשלוש קבוצות ראשיות, ששתיים מהן מוכרות גם היום: החיידקים (Bacteria) והארכאונים (Archaea). מהקבוצה
השלישית, האורקריוטים (Urkaryotes), התפתחו כל היצורים האיקריוטיים.
בתחילה, החיידקים הראשונים צרכו את האנרגיה הדרושה לקיומם ולהתפתחותם מתהליכים כימיים משחררי אנרגיה (חמצון וחיזור של תרכובות). אך כמות
האנרגיה הזאת היתה מוגבלת.
פריצת הדרך ה"אנרגטית", שאפשרה את האבולוציה של מגוון היצורים, היתה הופעתה של מערכת היכולה לקלוט את אנרגיית השמש ולהמירה לאנרגיה כימית.
רובנו מכירים את התהליך המכונה פוטוסינתזה, שבו נקלטת אנרגיית האור המגיעה מהשמש על-ידי הצבען הירוק כלורופיל, ומומרת לאנרגיה כימית.
תהליך זה מקורו בקבוצת החיידקים. גם בקבוצת הארכאונים הופיעה יכולת לקלוט את אנרגיית השמש, אך באופן שונה.
ב-1.5 מיליארד השנים הראשונות, החיידקים הפוטוסינתטיים, שבינתיים התפצלו לארבע קבוצות שונות, ביצעו פוטוסינתזה השונה מהפוטוסינתזה העיקרית
המוכרת לנו היום - היא לא פלטה חמצן, ולכן היא מכונה פוטוסינתזה שאינה חמצנית. לפני כשני מיליארדי שנה הופיעה הקבוצה החמישית - החיידקים
הכחוליים. קבוצה זו הצליחה לשלב בין שתי מערכות פוטוסינתטיות (photosystems) ממקורות שונים, וכתוצאה מכך הפוטוסינתזה מתבצעת תוך ביקוע של
מולקולות מים. התוצר של ביקוע שתי מולקולות מים היה מולקולת חמצן מולקולרי, O2, שנפלטה לאוויר (ביקוע כזה לא התאפשר בחיידקים בעלי מערכת
פוטוסינתטית אחת). לפיכך פוטוסינתזה זו מכונה פוטוסינתזה חמצנית. מאז עלה בהדרגה שיעור החמצן באוויר עד שהתייצב, לפני כ-800 מיליון שנה,
בריכוז המוכר לנו גם היום.
כתוצאה מעליית כמות החמצן באטמוספרה נוצרה שכבת האוזון, O3, שהיא מסננת יעילה של קרינה על-סגולה (קטלנית) המגיעה מהשמש.
נוכחות החמצן באוויר, ושכבת האוזון, היו שני תנאים הכרחיים להתפתחות מגוון החיים המוכר לנו היום.
אך החמצן הוא חומר רעיל ביותר לכל היצורים שאינם מותאמים אליו. הופעתו דחקה את מרבית המיקרואורגניזמים האל-אווירניים לאזורים ולמקומות
שחמצן אינו מגיע אליהם - כמעמקי הים, מעמקי האדמה ובתקופה מאוחרת יותר, מערכות עיכול של בעלי-חיים. בסביבה החמצנית שרדו רק שני טיפוסים
של מיקרואורגניזמים: אלה שהסתגלו לסביבה החמצנית ללא ניצול היתרונות שהיא מקנה, ואלה שהסתגלו לסביבה החדשה ולמדו לנצל את יתרונותיה.
היתרון העיקרי של סביבה חמצנית הוא אנרגטי. הראשון שפענח את היתרון הזה היה לואי פסטר (Pasteur): הוא גידל שמרים בנוכחות חמצן ובהיעדרו,
והראה שהצריכה של סוכר כגון גלוקוז (סוכר ענבים) קטנה בצורה משמעותית בתנאים אווירניים. תופעה זו נקראת עד היום על שמו - תוצא פסטר. אם
נפרוט את התופעה למטבעות אנרגיה (כמולקולות ה-ATP בתאים, המתווכות בהעברת האנרגיה בתא), הרי שבפירוק אל-אווירני של מולקולת גלוקוז מתקבלות
שתי מולקולות של ATP ואילו בפירוק חמצני של גלוקוז מתקבלות כ-30 מולקולות של ATP. לפיכך, בתנאים אווירניים התא צריך לנצל הרבה פחות גלוקוז
כדי להפיק אותה כמות של אנרגיה.
האיקריוטים, התאים בעלי הגרעין, הופיעו כנראה לראשונה לפני מיליארד וחצי שנים, כאשר האטמוספרה כבר היתה חמצנית, אם כי הכילה רק כ-10%
חמצן. כמעט כל התאים האיקריוטים מכילים מיטוכונדריונים, בניגוד לתאי החיידקים, והתעורר העניין בשאלה מה מקורם.
היתה זו החוקרת לין מרגוליס (Margulis) שהעלתה מחדש לסדר היום המדעי, בשנות השישים של המאה הקודמת, את התיאוריה האנדוסימביונטית. תאוריה מהפכנית זו הוצעה לראשונה ב-1883 על ידי הבוטנאי הצרפתי אנדראה שימפר (Schimper), שחקר את התחלקות הכלורופלסטים, ובעקבותיו, ב-1905, גם על ידי הבוטנאי הרוסי קונסטנטין מרצ'קובסקי (Mereschkowski), אך מלבד איזכורים בודדים פה ושם, כמעט ונשכחה.

למעלה, אנדראה שימפר (1856-1901); למטה, קונסטנטין מרצ'קובסקי (1855-1921)

התאוריה, המזוהה עד היום עם מרגוליס, שהתעקשה על אמיתותה למרות התנגדות הממסד המדעי, טוענת שהמיטוכונדריונים הם צאצאים של חיידקים אווירניים (שמסוגלים להשתמש בחמצן) שנבלעו או חדרו בדרך אחרת אל תוך תא איקריוטי ראשוני
(אורקריוט). חיידקים אלה יצרו סימביוזה שהפכה הכרחית עם התא המאכסן שלהם. סימביוזה זו נתנה יתרון עצום למאכסן - הוא יכול היה להפיק הרבה
יותר אנרגיה מחומרי המזון שבסביבתו. התיאוריה מתייחסת גם לכלורופלסטים - אותם אברונים תוך-תאיים באצות ובצמחים, המבצעים את
הפוטוסינתזה החמצנית. וטוענת שגם הם חיידקים שנבלעו - חיידקים כחוליים.

מה היו העדויות שתמכו בתיאוריה האנדוסימביונטית? גם המיטוכונדריון וגם הכלורופלסט הם יחידות אוטונומיות בתוך התא
האיקריוטי. גודלם דומה מאוד לזה של חיידקים. כל אחד מהם עטוף בממברנה (קרומית) כפולה, כמו חיידק, ויש להם חומר תורשתי (DNA) נפרד ומעגלי,
כמו בחיידקים. יש בהם ריבוזומים (אתרי ייצור חלבונים) השונים במבנם מהריבוזומים הציטופלזמיים, ודומים במבנם לריבוזומים חיידקיים. גם
תרופות אנטיביוטיות הפוגעות בחיידקים, ושאינן פוגעות בתאים איקריוטיים, מעכבות את פעילות המיטוכונדריונים והכלורופלסטים, כאשר הם מבודדים.
כמו כל תיאוריה מהפכנית, גם תיאוריה זו לא התקבלה לאלתר. אך בעקבות פיתוח השיטה לקביעת הרצפים של ה-DNA, החלה להתפתח השיטה החדשה למיון
היצורים המבוססת על השוואת רצפי DNA של גנים שמורים (כלומר גנים שאינם מאפיינים קבוצה מצומצמת של יצורים, אלא קיימים בכל עולם החי). שיטה
זו אפשרה להוכיח שאכן התיאוריה נכונה. המיטוכונדריון מקורו בקבוצה של חיידקים אווירניים מקבוצת הפרוטאובקטריה, ואילו מקורם של הכלורופלסטים
הוא בחיידקים הכחוליים.
ולא זו בלבד: החיידקים הקרובים ביותר למיטוכונדריון המוכרים לנו היום הם הריקֶציות (Rickettsia): אלה הם חיידקים קטנים, המהווים טפילים
תוך-תאיים וגורמים למחלות כמו טיפוס (epidemic thyphus) וקדחת הבהרות (spotted fever). גם הם, כמו המיטוכונדריון, אינם מסוגלים להתקיים
מחוץ לתא המאכסן. אנו משערים שיחסים טפיליים של אבי המיטוכונדריון עם המאכסן שלו השתנו ליחסי גומלין שהביאו להתפתחותו של התא האיקריוטי
החמצני, שעם צאצאיו נמנים גם אנחנו.

החיפוש אחר המאכסן המקורי של אבי המיטוכונדריון (תא איקריוטי ללא מיטוכונדריה) לא צלח. אנו מניחים שהתא החדש, הודות ליתרון האנרגטי
שרכש, לא השאיר סיכוי למתחריו בסביבה החמצנית המתהווה, ואלה נכחדו. אמנם קיימים היום כמה יצורים איקריוטיים חסרי מיטוכונדריונים, החיים
בסביבות אל-אווירניות (כגון טפיל המעיים Giardia lamblia) - אך חיפוש מדוקדק ב-DNA שלהם מגלה לנו שהם צאצאים של יצורים בעלי
מיטוכונדריונים, שאיבדו את המיטוכונדריונים שלהם.
ומה עם הכלורופלסטים? כיום מקובל להניח, שתא איקריוטי (בעל מיטוכונדריונים) בלע בשלב מאוחר יותר חיידק כחולי ויצר איתו סימביוזה הכרחית
שהובילה להתפתחות האצות והצמחים. אך בעוד שאירוע הסימביוזה עם אבי המיטוכונדריון היה כנראה, חד-פעמי, נראה שהיו שלושה אירועים שונים של
יצירת סימביוזה עם חיידק כחולי. מאחד מהם נוצרו, כאמור, האצות הירוקות והצמחים, ומשני האחרים שתי קבוצות אחרות של אצות.
שנים רבות של אבולוציה משותפת בין התאים האיקריוטיים והמיטוכונדריונים שלהם גרמו למעבר של גנים מהמיטוכונדריון לתא המאכסן ולהפך,
והמיטוכונדריון מעורב בתהליכים תאיים נוספים, שאחד מהם הוא ההתאבדות של תאים (אפופטוזה).
חשוב לזכור שביצורים כמונו, בעלי רבייה מינית, המיטוכונדריונים עוברים בדרך-כלל בתורשה אמהית - לאחר ההפריה, בתא הביצה יש מאות
מיטוכונדריונים שמקורם בתא הביצה ורק בודדים שמקורם בתא הזרע. (המיטוכונדריונים שמקורם בתא הזרע מסומנים מראש להשמדה לאחר ההפריה על-ידי
יוביקוויטין - ראו "חלבונים ומפרקים", "גליליאו"
38.) לפיכך, בבדיקת אמהוּת ניתן להסתפק בבדיקת DNA מיטוכונדרי בלבד. עם זאת, יש להביא בחשבון שבתדירות נמוכה מאוד, יש גם הורשה אבהית של
מיטוכונדריונים.
בכל אחד מתאי גופנו שוכנים, אם כן, כ-50 צאצאי חיידקים, המשתפים פעולה עם התא ואחראים ליכולתו לקיים מטבוליזם חמצני. חיידקים אחרים,
כחוליים, וצאצאיהם הכלורופלסטים, דואגים לאספקת החמצן שמנצלים שאר היצורים על פני כוכב-הלכת שלנו.





,_1887_Wellcome_L0062031.jpg)


















אין תגובות:
הוסף רשומת תגובה